Результаты адаптации молоди судака к выращиванию в индустриальных условиях
Судак Sander lucioperca (Linnaeus, 1758) [1] – ценная промысловая хищная рыба семейства окуневых. Является одним из основных пресноводных промысловых видов рыб. Однако в последние десятилетия под влиянием антропогенных факторов запасы судака значительно сократились и находятся в депрессивном состоянии [2, 3].
В Азовском море интенсивность изъятия судака промыслом в первой половине XX в. находилась на уровне 60 % от численности популяции в год. Тем не менее запас, а соответственно, и уловы, оставались высокими. При ослаблении промысла, даже кратковременном, численность судака значительно увеличивалась. Столь высокая пластичность популяции судака, служившая основой высокоинтенсивного, но стабильного промысла, обеспечивалась главным образом двумя факторами. Во-первых, наличием большого количества высокопродуктивных нерестилищ, общая площадь которых, по подсчетам разных авторов, достигала 5–7 тыс. км2 [4].
Во-вторых, низкой соленостью Азовского моря (в среднем 9,5–10,5 0/00), что позволяло судаку осваивать кормовые объекты практически на всей его акватории, включая Таганрогский залив.
Оба фактора во многом зависят от гидрологического режима рек, впадающих в Азовское море.
Поэтому антропогенное преобразование стока двух наиболее крупных рек Азовского бассейна – Дона и Кубани – не могло не оказать влияния на всю экосистему Азовского моря, в том числе и на состояние популяции полупроходного судака.
Изменение гидрологических условий и последовавшие за ними гидробиологические преобразования оказали влияние на ихтиофауну Азовского моря в целом [5, 6], в том числе на популяцию полупроходного судака [7]. Наблюдается неуклонное сокращение запасов, снижение эффективности естественного и искусственного воспроизводства судака. Так, если в 1930-е гг. уловы судака в Азовском бассейне достигали 74 тыс. т, то к 1990–2004 гг. они снизились до 1–3 тыс. т. В Волго-Каспийском бассейне запасы судака находятся в критическом состоянии, его вылов в 2005–2006 гг. составлял 0,38–0,42 тыс. т, что в 5 раз меньше, чем в начале 1990-х гг. Численность молоди судака на нерестилищах дельты Волги в последние годы очень низкая, в 2005 г. она не превысила 0,4 тыс. экз./га [3]. Произошло снижение таких показателей, как средняя масса и длина рыб. Сокращение в структуре стада особей старшего возраста свидетельствует о сохранении чрезмерного антропогенного давления на популяцию [8].
Воспроизводство волжского проходного судака не зависит от состояния кормовой базы, величина его запасов определяется масштабами и эффективностью размножения. В годы с низким объемом весеннего половодья в дельте Волги она снижается катастрофически, и единственным способом поддержания запасов судака является заводское разведение в достаточно широких масштабах [9, 10].
Работы по разведению полупроходного судака в дельте Волги были начаты в конце 40-х гг. XX столетия. На первом этапе (до конца 70-х гг.) для этой цели использовались нерестово-выростные хозяйства (НВХ). В одном водоеме совмещались процессы нереста, инкубации икры и выращивания молоди нескольких видов рыб: леща и судака; сазана и судака; сазана, леща и судака [11]. Эффективность этой биотехнической схемы была невелика: выход покатной молоди составлял не более 20–25 тыс. шт./га, или 0,5–5 тыс. шт. на одну самку, максимальная выживаемость от икры до покатной молоди не превышала 5 %. Выпуск молоди судака в 50–60-е гг. (по данным ФГУ «Севкаспрыбвод») составлял в среднем 35–66 млн экз. в год; во второй половине 70-х гг., в связи с резким снижением уловов рыб в дельте и невозможностью заготовить производителей весной, он уменьшился до 12 млн экз., а затем был прекращен.
В 60-е гг. в экспериментальных условиях были разработаны основы биотехники заводского разведения судака; процессы нереста, инкубации икры и выращивания молоди были разделены, благодаря чему они стали более управляемыми [12].
Эта схема, по сравнению с биотехникой разведения молоди в НВХ в экспериментальных условиях, позволила повысить выживаемость от икры до покатной молоди с 5 до 15 %, выход молоди с одного га выростной площади – с 20–25 до 100 тыс. шт. и, следовательно, сократить количество производителей в 3, а выростные площади – в 5 раз.
В настоящее время основным способом поддержания запасов судака является искусственное воспроизводство, которое характеризуется небольшими масштабами: выпуск молоди в 1998–2002 гг. колебался от 1,7 млн шт. до 10 млн шт., составив в среднем 5,1 млн шт. Увеличение объемов выпуска молоди ограничивается проблемой заготовки производителей [13]. Для пополнения численности и поддержания запасов судака на достаточно высоком уровне необходимо разработать биотехнику индустриального выращивания этого ценного вида в полностью контролируемых условиях [14].
Приспособляемость рыб к питанию определенными кормами непостоянна и меняется по мере их роста, изменения строения ротовой полости и пищеварительной системы. Смена кормов в онтогенезе позволяет виду осваивать различные корма, и при этом у большинства рыб по мере их перехода с одной стадии развития на другую наблюдается расширение спектра питания – увеличение количества компонентов пищи. Успех же разработки эффективных и экологически безопасных кормов зависит от знания пищевых потребностей рыб, которые подвержены влиянию целого ряда факторов (включая абиотические).
Имеющиеся данные свидетельствуют о том, что ферменты, обеспечивающие процессы пищеварения у рыб, адаптированы к спектру и интенсивности их питания, а также к составу пищи. Обнаруженная пластичность ферментных систем играет важную роль в приспособлении рыб, большинство из которых являются эврифагами, к условиям жизнедеятельности и питания. Поэтому вопросы адаптации рыб к новым индустриальным методам содержания являются очень актуальными при выращивании объектов в условиях замкнутого водоснабжения.
В связи с вышеизложенным целью исследований явилась разработка методов адаптации судака к индустриальным условиям выращивания.
Материал и методы исследований
Исследования по адаптации молоди судака к индустриальным условиям выращивания проводили в экспериментальном аквариальном комплексе базовой кафедры Южного научного центра РАН «Аквакультура и водные биоресурсы» Астраханского государственного технического университета. Прудовая молодь судака массой 1,5 г была завезена в июне 2008 г. Содержание и выращивание молоди осуществляли в стеклопластиковом лотке площадью 2,8 м2 с замкнутым циклом водоснабжения. В период экспериментальных работ гидрохимические показатели воды поддерживали на оптимальном уровне. Температура воды при проведении экспериментов составляла 20,5–21,5 °С, содержание O2 – 7,8–8,4 мг/л, рН – 7,3–7,5.
Оценку поведенческих реакций молоди проводили в тесте «открытое поле». Рыб по одной помещали в камеру размерами 60 ? 60 см со слоем воды около 5 см во избежание значительных перемещений рыбы. На дно камеры была нанесена координатная сетка с размерами квадратов 10 ? 10 см. Удвоенное количество пересечений координатных линий за первые 30 секунд после помещения рыбы в новую обстановку характеризует ориентировочную активность молоди (ОА, ед./мин). Усредненное количество пересечений координатных линий рыбой за период с 1-й по 7-ю минуту опыта расценивается как фоновая двигательная активность (ФА, ед./мин).
Через 7 минуту после начала эксперимента применяли раздражитель, имитирующий нападение хищника. Удвоенную двигательную активность тестируемой молоди в течение 3 минут после воздействия этого стимула расценивали как реактивность на визуально-динамический стимул, или реакцию на хищника (РА1 хищн., ед./мин). Через 8,5 минуту после начала эксперимента применяли первый виброакустический раздражитель – высокочастотный удар (ВЧ). Удвоенное количество пересечений рыбой сторон квадратов за первые 30 секунд после нанесения удара определяли как реактивность на виброакустический стимул (РА2 вибр., ед./мин). Через 10 минут был применен следующий раздражитель – световая вспышка длительностью 4 секунды и освещенностью 5 000 люкс. Удвоенная величина двигательной активности за первые 30 секунд после предъявления светового раздражителя определялась как реактивность на третий сигнал – (РА3, ед./мин). На 11,5 минуте применяли второй виброакустический раздражитель: удар молоточком по стенке камеры (глухой низкочастотный звук (НЧ)) и регистрировали реактивность объекта исследования на 4-й сигнал – (РА4, ед./мин). Для оценки двигательной активности использовали 10 шт. молоди судака [15].
Контроль за темпом роста осуществляли один раз за 10–15 суток. Взвешивание и измерение рыбы проводили согласно рекомендациям И. Ф. Правдина [16]. Контрольным промерам подвергали всю рыбу, находящуюся в эксперименте.
Опыты проводили в двукратной повторности, данные подвергали статистической обработке по Г. Ф. Лакину [15] с применением персонального компьютера. При этом использовали элементы статистического анализа с определением средней X и среднего отклонения а. Сравнительные признаки оценивали с помощью критерия Стьюдента. Каждый из вариантов сопоставляли с другими, причем разность принимали достоверной при первой степени вероятности безошибочного суждения (р > 0,05).
Результаты исследований
В настоящее время существует проблема с заготовкой необходимого количества производителей судака, влекущая за собой проблему формирования ремонтно-маточных стад судака в искусственных условиях. Это возможно путем адаптации к индустриальным условиям разновозрастного судака, заготовленного из естественных условий, и заводской молоди судака, полученной в искусственных условиях «от икры».
При выращивании судака используют в основном прудовый метод выращивания. Молодь содержится в прудах на естественных кормах, поэтому после посадки в бассейн она будет испытывать стресс, связанный с изменениями условий обитания.
В связи с этим разработка методов первичной адаптации, оптимальных параметров содержания и рациональных методов кормления данного объекта в индустриальных условиях является весьма актуальной.
В течение первых 4–6 часов после транспортировки осуществляли адаптацию прудовой молоди судака к температурным условиям, постепенно выравнивая температуру. В первые трое суток кормление молоди не проводили, стараясь не беспокоить рыбу. После первичной адаптации на 4-е сутки начали кормление мотылем и предварительно измельченными дождевыми червями. Молодь активно потребляла корм. По мере роста молоди данный вид корма постепенно был заменен на фарш из кильки и частиковых видов рыб. На 16-е сутки рацион полно- стью состоял из рыбного фарша. Кратность кормления составляла 3–4 раза в сутки. Остатки корма удаляли через 30 минут после кормления. На 38-е сутки выращивания в корм стали вводить гранулированные комбикорма.
Рыбоводные показатели выращивания молоди судака представлены в таблице. За период исследований абсолютный прирост молоди составил 3,0 г, среднесуточный – 0,05 г (табл.).
Рыбоводно-биологические показатели молоди судака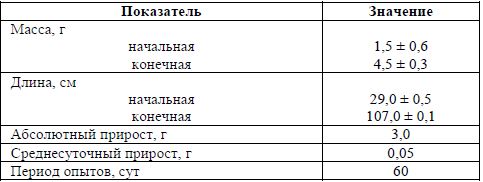
От условий выращивания гидробионтов зависит их физиологическое состояние, которое отражает уровень развития адаптационной системы. Применяя определенные методики исследований, можно сделать вывод о физиологическом статусе организма и степени воздействия условий обитания рыб [14].
Особого внимания заслуживают условно-рефлекторные методики оценки способности рыб к приобретению и сохранению поведенческих навыков. Поскольку в искусственных и естественных водоемах экологические условия резко различаются, крайне важной является быстрая выработка молодью в индустриальных условиях приспособительных реакций, необходимых для выживания в новой среде обитания.
Известно, что в раннем онтогенезе развитие организма зависит как от внешних факторов среды, так и от взаимодействия внутренних систем. Согласно теории Анохина, основополагающим внутренним фактором является рецепторный комплекс, осуществляющий связь организма с внешней средой. Развитие высшей нервной деятельности, которая способствует развитию и функционированию систем организма и, следовательно, согласованной деятельности внутренних органов, находятся в прямой зависимости от степени развития всех рецепторных систем.
Совершенно очевидно, что условия содержания будут являться определяющими при развитии всех систем организма.
Поэтому было необходимо проверить у молоди судака изменение реактивности высшей нервной деятельности как показателя адаптационных способностей организма в раннем онтогенезе при содержании в бассейнах.
Для оценки физиологического состояния молоди судака были проведены испытания в тесте «открытое поле». Результаты теста представлены на рис. 1.

Рис. 1. Результаты тестирования молоди судака
Ориентировочная активность является показателем реактивности центральной нервной системы. В естественных условиях в результате низкой двигательной активности при попадании в незнакомые условия молодь становится доступной хищникам [17]. Молодь судака, содержащаяся в искусственных условиях, при попадании в незнакомую обстановку демонстрировала реакцию затаивания, переходящую в медленные перемещения по опытной установке.
Из рис. 1 видно, что реакция на новую обстановку у молоди судака заторможена, период затаивания затягивается, поисковая реакция наступает значительно позже. При содержании молоди в искусственных условиях в течение 2-х недель произошло изменение ОА. Так, в начале показатель в среднем составил 2,3 ± 0,61, а затем повысился до 2,56 ± 1,18.
Фоновая активность молоди судака в начале выращивания была значительно выше ОА.
В этот период рыба сравнительно быстрее передвигалась по экспериментальной установке, проявляя поисковые реакции. В последующем молодь стала проявлять меньшую подвижность.
Через две недели значение ФА снизилось в 2 раза – с 3,08 ± 0,32 до 1,25 ± 0,26 ед., что свидетельствует об адаптации рыб к искусственным условиям.
При применении первого раздражителя (РА1 хищн.) рыба проявляла реакцию затаивания, что является характерным для судака, т. к. он сам ведет хищнический образ жизни и при малейшей опасности затаивается. В начале выращивания этот показатель был значительно выше – 2,5 ± 0,53.
Второй раздражитель (РА2) – высокочастотный удар – относится к адекватным биотическим раздражителям, имитирующим звуки питания хищных видов рыб, характеризует степень развития акустической системы организма в раннем онтогенезе. Наиболее выраженная реакция затаивания молоди проявилась в начальный период – 3,1 ± 0,52, затем она снизилась в 1,6 раза и составила 1,9 ± 0,52 ед. При сравнении реакции на раздражитель и после его действия в течение минуты и в первом, и во втором случае различия достоверны при t > 0,05.
Реакция молоди на данный раздражитель вначале демонстрировала более динамичный характер, молодь затаивалась и ее двигательная активность характеризовалась бросковым поведением. При втором тестировании затаивание носило кратковременный характер, после чего рыбы совершали медленные плавательные движения.
Третий раздражитель (РА3) – вспышка яркого света – неадекватный биотический фактор, характеризует имитацию увеличения освещенности, при которой возникает пищедобывательная реакция [18]. По сравнению с предыдущей испуговой реакцией молодь снизила двигательную активность. Реакция сохранялась на протяжении всего времени действия света. Этот раздражитель, в отличие от предыдущих, носит сигнальный характер. И на этот сигнал рыбы проявляли достоверно пониженную двигательную активность.
Четвертый раздражитель (РА4) – низкочастотный удар. В естественных условиях низкочастотные раздражители являются естественным шумом реки. Реакция молоди на этот раздражитель повысилась по сравнению с реакцией на вспышку яркого света. Так, в первом случае она составила 2,8 ± 0,6, во втором – 1,7 ± 0,47 ед. Это свидетельствует о более выраженной реактивности рыб на адекватные раздражители при попадании в естественную среду. Именно в этот период адаптивные качества молоди определяют ее выживаемость.
Таким образом, тестирование молоди по всем раздражителям показало, что в начале выращивания все параметры, кроме ориентировочной активности были значительно выше.
Со временем реактивность молоди судака понизилась, что связано с ее адаптацией к условиям содержания. Не произошло повышения реагирования на высоко- и низкочастотные удары, что, возможно, объясняется тем, что молодь содержалась в информационно обедненной среде в отличие от естественного водоема. Молодь судака в раннем онтогенезе выращивалась в прудовых условиях, происходило формирование рецепторного комплекса. При помещении в несвойственную среду – искусственные условия – произошло торможение в ее развитии. Чтобы избежать этого, необходимо создание естественного фона среды: течение с запахом пищевых объектов скоростью от 0,1 до 1–2 м/с; естественная освещенность с соблюдением фотопериода; для развития акустико-латеральной системы – создание фона, соответствующего естественным адекватным частотам в диапазонах 30–120, 250–500, 900–1 600 Гц.
Разработанная схема адаптации представлена на рис. 2.

Рис. 2. Схема первичной адаптации
Выращивание рыб в индустриальных условиях всегда сопряжено с существенным изменением их физиологического состояния. Использование высококачественных кормов является обязательным условием эффективного выращивания рыб. В противном случае возможно развитие алиментарных болезней, которое часто сопровождается вторичными инвазиями, в частности протозойными и грибковыми. При адаптации молоди судака, из-за несвойственных ей условий обитания, происходило нарушение обмена веществ (жировая дистрофия печени), поэтому адаптация к искусственному корму требует дальнейшего изучения.
Разработка методики адаптации судака к индустриальным условиям позволит сформировать ремонтно-маточное стадо для целей его воспроизводства и товарного выращивания.
Заключение
По результатам первых опытов разработаны методы адаптации судака к искусственным условиям, которые составляют первый этап приучения рыб к искусственным условиям. Особого внимания заслуживают условно-рефлекторные методики оценки способности рыб к приобретению и сохранению поведенческих навыков. Поскольку экологические условия в искусственных и естественных водоемах резко различаются, крайне важной является быстрая выработка молодью в индустриальных условиях приспособительных реакций, необходимых для выживания в новой среде обитания.
Исследования показали снижение реактивности молоди судака по мере ее адаптации к условиям содержания. Не произошло повышения реагирования на высоко- и низкочастотные удары, поскольку молодь содержалась в информационно обедненной среде в отличие от молоди естественного водоема.
М. Н. Сорокина, А. В. Ковалёва, Е. Н. Пономарёва